
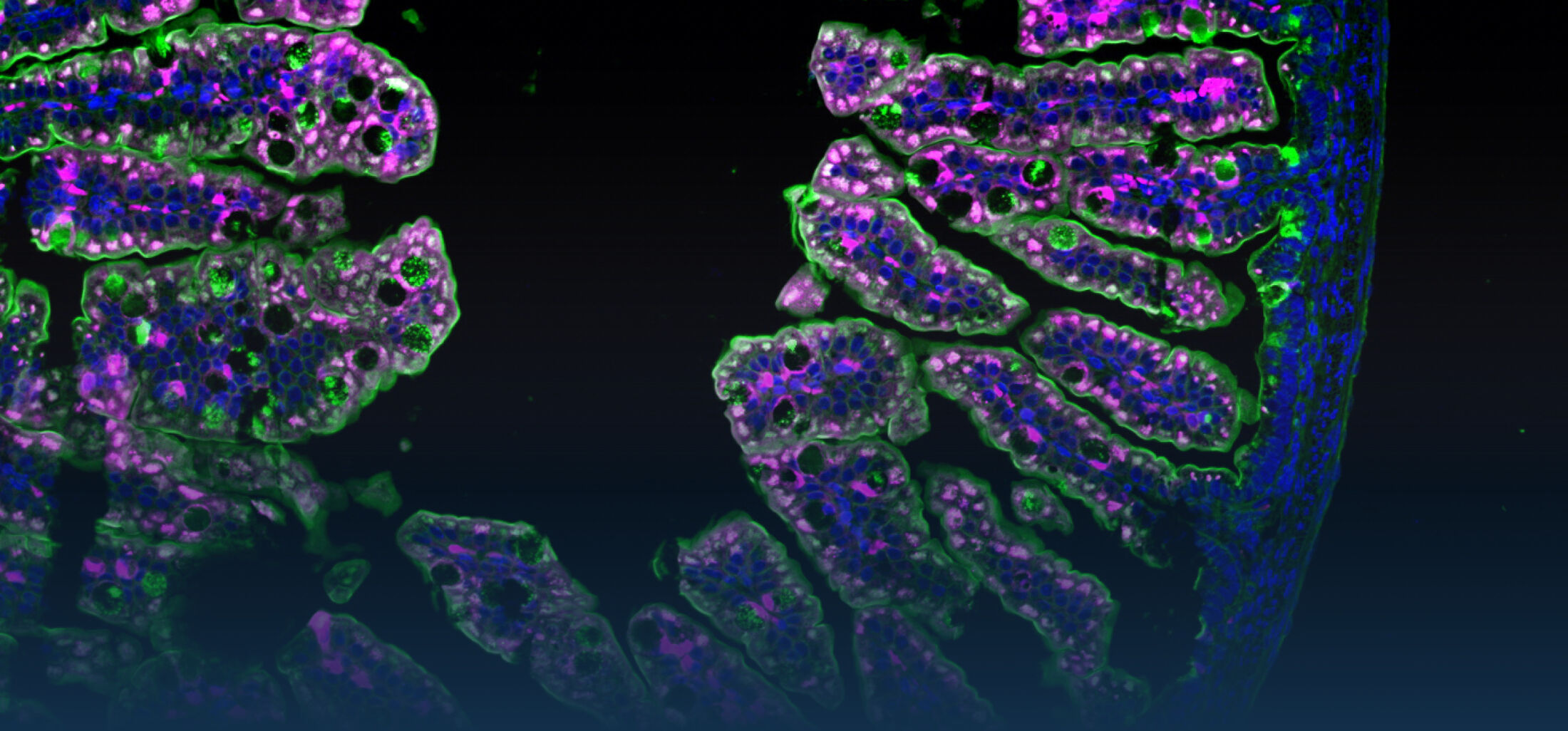
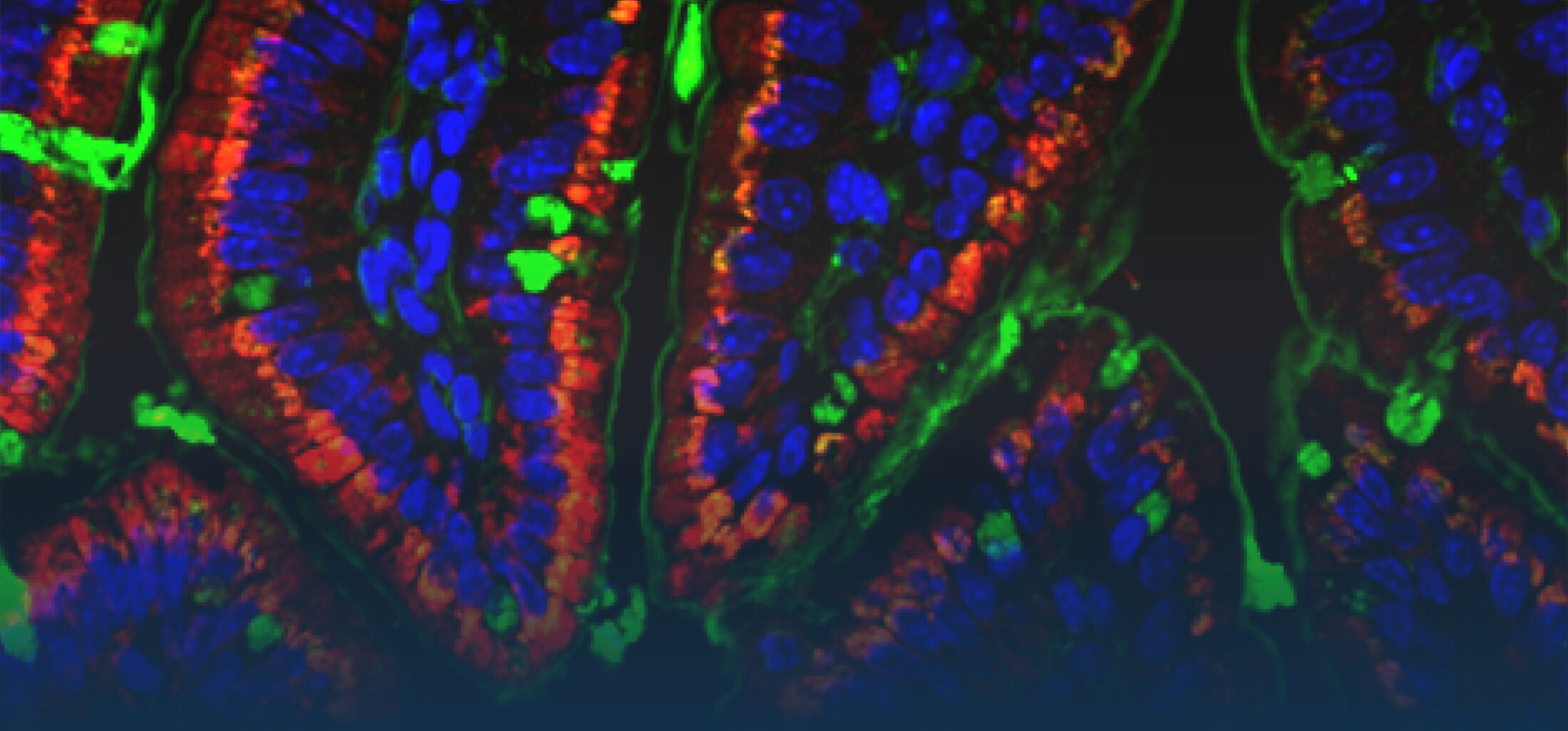



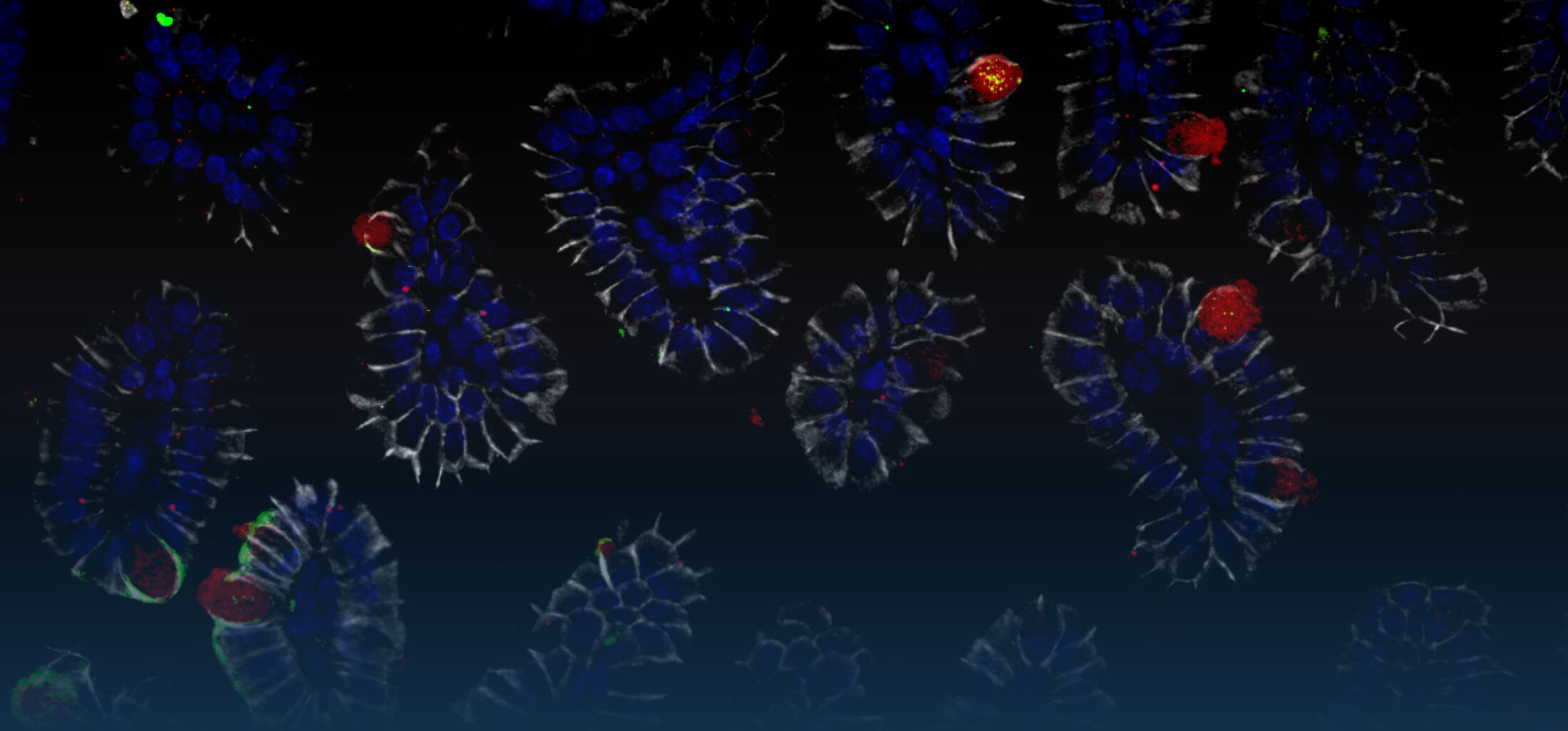















DIE ROLLE DES ANGEBORENEN IMMUNSYSTEMS IM INTESTINALTRAKT
1.DIE ERKENNUNG KOMMENSALER UND ENTEROPATHOGENER ERREGER DURCH EPITHELIALE REZEPTOREN DES ANGEBORENEN IMMUNSYSTEMS
Rezeptoren des angeborenen Immunsystems ermöglichen die Erkennung von konservierten molekularen Strukturen von Bakterien, Pilzen, Viren und Parasiten. Eine Stimulation der Rezeptoren induziert die rasche Aktivierung des Immunsystems z.B. durch die Sekretion von Chemokinen und Zytokinen und damit die Rekrutierung von professionellen Phagozyten. Im Fall einer lokalen Infektion kann dies zur frühzeitigen Elimination des Erregers führen und ein weiteres Fortschreiten der Erregerausbreitung im Wirt verhindern. Eine überschießende unkontrollierte Stimulation des angeborenen Immunsystems oder eine Aktivierung in der Abwesenheit eines Infektionserregers kann, auf der anderen Seite, zu chronischen Entzündung und damit zur Krankheitsentstehung beitragen.
Ziel unserer Arbeiten ist ein besseres Verständnis der Funktion, Bedeutung und Regulation von Rezeptoren des angeborenen Immunsystems auf Darm-epithelzellen. Epithelzellen bilden die oberste Zellschicht der Darmschleimhaut, die beim erwachsenen Wirtsorganismus durch eine dichte und vielfältige bakterielle Flora (Mikrobiota) besiedelt ist. Die anatomische Lokalisation der Epithelzellen an der Grenzfläche zwischen sterilem Wirtsgewebe und dem bakteriell besiedeltem und mit Nahrungsbestandteilen exponiertem Darmlumen erfordert eine strikte Kontrolle der Stimulation von Immunrezeptoren, um eine ungewollte, gewebezerstörende Reaktion des Immunsystems zu verhindern. Auf der anderen Seite ist der Darm das Ziel vieler pathogener Mikroorganismen; eine frühzeitige Erkennung und Elimination kann deshalb zur Vermeidung von Infektionen beitragen.
Schon früh konnten wir die Expression von Toll-like Rezeptor (TLR)4, dem Rezeptor für bakterielles Lipopolysaccharid (LPS) durch Darmepithelzellen zeigen (Hornef et al., 2002) und Besonderheiten der zellulären Lokalisation für die Ligandenerkennung charakterisieren (Hornef et al., 2003; Duerr et al., 2009). Auf der Suche nach regulierenden Mechanismen entdeckten wir, dass Darmepithelzellen kurz nach Geburt einen adaptiven Prozess durchlaufen, der die Aktivierung von TLRs während der ersten Tage nach Geburt hemmt und damit eine ungewollte Stimulation während der postnatalen bakteriellen Kolonisierung der Darmmukosa verhindert (Lotz et al., 2006). Eine microRNA-vermittelte Translations-inhibition sowie die proteosomale Degradation des Signalmoleküls interleukin 1 receptor associated kinase (IRAK)1 verursachen diese adaptive Tolerisierung des Darmepithels nach Geburt und
schützen es vor einer ungewollten Immunstimulation während der Etablierung
der Darmflora (Chassin et al., 2010). Beim gesunden Erwachsenen trägt hingegen eine kontrollierte Stimulation von epithelialen TLRs zur Aufrechterhaltung der Darmhomöostase bei (Stockinger et al., 2014). Jedoch findet sich im adulten Wirt nach einer Ischämie durch SUMOylierung und einen Wechsel der Ubiquitinierung (K48 zu K63) eine vermehrte epitheliale
IRAK1 Expression. Die erhöhten IRAK1 Spiegel vermitteln eine postischämische
Überempfindlichkeit gegenüber mikrobiellen Liganden und tragen zur
Gewebeschädigung nach transienter Gewebehypoxie bei (Chassin et al., EMBO
Mol Med, 2012). IRAK1 spielt eine wichtige Rolle bei der Signaltransduktion bei vielen TLRs. Einzig TLR3, der Rezeptor für doppelsträngige RNA, induzierte eine IRAK1 unabhängige Signalkaskade. Interessanterweise findet sich im neonatalen Darmepithel eine stark reduzierte Expression von TLR3. Diese reduzierte TLR3 Expression trägt dazu bei, dass der neugeborene Wirt gegenüber einer Infektion mit Rotaviren eine deutlich erhöhte Empfindlichkeit zeigt (Pott et al., 2012). TLR3 kooperiert mit den Helikasen RigI und MDA-5 bei der Erkennung von pathogenen dsRNA Viren. Diese Rezeptoren induzieren die Sekretion von Interferonen, die Zellen gegenüber einer Virusinfektion unempfindlich machen. Bei Untersuchungen zur epithelialen Immunantwort
auf eine Rotavirusinfektion konnten wir erstmals die besondere Bedeutung von Interferon-λ für die antivirale Wirtsantwort des Darmepithels
zeigen (Pott et al., 2011). Eine Infektion mit Listeria monozytogenes stimuliert nach Lyse der endosomalen Membran in Darmepithelzellen Nod2, den im Zytosol lokalisierten Rezeptor für Muramyl Dipeptid (MDP) aus der bakteriellen Peptidoglycanschicht. Mithilfe Einzelzellanalyse konnten wir die überraschende Beobachtung machen, dass trotz der Invasions-abhängigen Aktivierung
von Darmepithelzellen v.a. nicht infizierte, Listeria-negative Epithelzellen Chemokine produzierten. Weitere Analysen bestätigten die Kommunikation zwischen Darmepithelzellen mittels durch die NADPH Oxidase (NOX)4 produzierten Sauerstoffradikalen. Damit konnte ein neuer Mechanismus der
Informationsweitergabe an benachbarte Darmepithelzellen nach bakterieller
Infektion gezeigt werden (Dolowschiak etal., 2010).
Eine orale Exposition Neugeborener mit enteropathogenen Bakterien wie z.B.
Salmonella enterica führt wegen der niedrigen Diversität der Darmflora zu einer
raschen Kolonisierung des Darmes. Im weiteren Verlauf findet sich im neugeborenen Wirt überraschenderweise eine Salmonella pathogenicity island
(SPI)1-abhängige Invasion von Darmepithelzellen und die Ausbildung von
intraepithelialen Mikrokolonien. Diese Strukturen wurden zuvor v.a. nach
Infektion von Zellkulturen beobachtet. Mit diesen Arbeiten konnten wir erstmals ein leicht zugängliches in vivo Modell zur Analyse der Salmonella/Epithelzell-
Interaktion etablieren (Zhang et al., 2014).
Hornef MW, Normark BH, Vandewalle A, Normark S. Intracellular recognition of
lipopolysaccharide by toll-like receptor 4 in intestinal epithelial cells. J Exp Med. 2003; 198(8): 1225-35.
Hornef MW, Frisan T, Vandewalle A, Normark S,Richter-Dahlfors A. Toll-like receptor 4 resides in the Golgi apparatus and colocalizes with internalized
lipopolysaccharide in intestinal epithelial cells. J Exp Med. 2002; 195(5): 559-70.
Lotz, M., Gütle, D., Ménard, S., Walther, S.,Bogdan, C., Hornef, M.W.. Postnatal acquisition of endotoxin tolerance in intestinal epithelial cells. J Exp
Med. 2006; 203(4): 973-84.
Duerr CU, Zenk SF, Chassin C, Pott J, Gütle D, Hensel M, Hornef MW. O-antigen delays lipopolysaccharide recognition and impairs antibacterial host defense in murine intestinal epithelial cells. PLoS Pathog. 2009 ;5(9): e1000567.
Chassin, C., Kocur, M., Pott, J., Duerr, C.U., Gütle, D., Lotz, M., Hornef, M.W. miR-146a mediates protective innate immune tolerance in the neonate intestine. Cell Host Microbe 2010, 8: 358-68.
Dolowschiak, T., Chassin, C., Ben Mkaddem, S., Fuchs, T.M., Weiss, S., Vandewalle, A., Hornef, M.W.. Potentiation of epithelial innate host responses by intercellular communication. PLoS Pathogens 6: e1001194,2010.
Pott, J., Mahlakõiv, T., Mordstein, M., Duerr, C.U., Michiels, T., Stockinger, S.*, Staeheli, P.*, Hornef, M.W.*. IFN- determines the intestinal epithelial antiviral host defense. Proc Natl Acad Sci U S A. 108: 7944-9, 2011.
Chassin C, Hempel C, Stockinger S, Dupont A, Kübler JF, Wedemeyer J, Vandewalle A, Hornef MW. MicroRNA-146a-mediated downregulation of IRAK1 protects mouse and human small intestine against ischemia/reperfusion injury. EMBO Mol Med. 4(12):1308-19, 2012
Pott J, Stockinger S, Torow N, Smoczek A, Lindner C, McInerney G, Bäckhed F, Baumann U, Pabst O, Bleich A, Hornef MW. Age-dependent TLR3 expression of the intestinal epithelium contributes to rotavirus susceptibility. PLoS Pathogens, 8(5):e1002670. 2012
Fulde, M., Hornef, M.W. Maturation of the mucosal innate immune system during the postnatal period. Rev Immunol. 260: 21-34, 2014.
Zhang, K., Dupont, A., Torow, N., Gohde, F., Leschner, S., Lienenklaus, S., Weiss, S., Brinkmann, M.M., Kühnel, M., Hensel, M., Fulde., M.*, Hornef, M.W. Age-dependent enterocyte invasion and microcolony formation by Salmonella. PLoS Pathogens, 2014 (in press)
Stockinger, S., Duerr, CU., Fulde,, M., Dolowschiak, T., Pott, J., Yang, I., Eibach, D., Bäckhed, F., Akira, S., Suerbaum, S., Brugman, M., Hornef, M.W. TRIF Signaling Drives Homeostatic Intestinal Epithelial Antimicrobial Peptide Expression. J. Immunol. 2014 (in press)
Torow, N., Yu, K., Hassani, K., Bleich, A., Lochner, M., Brenneke, A., Weiss, S., Förster, R., Pabst, O., Hornef, M.-W. Active suppression of intestinal CD4+TCRαβ+ T lymphocyte maturation during the postnatal period. (submitted).
2. FAKTOREN DER EPITHELIALEN IMMUNABWEHR: ENTERISCHE ANTIMIKROBIELLE PEPTIDE
Ein zweiter Schwerpunkt unserer Arbeiten liegt auf der Analyse der intestinalen epithelialen Wirtsabwehr. Im Zentrum unseres Interesses stehen dabei die Produktion und Regulation enterischer antimikrobieller Peptide. Paneth-Zellen am Grund intestinaler Krypten produzieren und sekretieren eine große Menge und Vielfalt enterischer antimikobieller Peptide wie -Defensine und cryptdin related sequence (CRS)-Peptide. Diese vom Körper selbst hergestellten Antibiotika-ähnlichen Substanzen scheinen eine wichtige Rolle bei der mukosalen Wirstabwehr, aber auch bei der Regulation der enterischen Mikrobiota zu spielen.
In weiteren Arbeiten konnten wir zeigen, das CRS Peptide sich aus Peptid-Homo- oder Heterodimeren zusammensetzten. Dieser Mechanismus erhöht die Variabilität des Peptid-Repertoires und könnte wichtig sein, um gegen alle potentiell gefährlichen Bakterien eine wirksame Abwehr bilden zu können (Hornef et al., 2004).
Die Sekretion von antimikrobiellen Peptiden durch Paneth Zellen wird durch neuronale Stimuli (Parasympatiko-mimetika) und mikrobielle Stimuli (MDP) induziert. Neuere Arbeiten an isolierten Darmkrypten identifizierten kürzlich auch endogene Mediatoren wie IL-4 und IL-13 als Stimuli für die Degranulation von Paneth Zellen und die Freisetzung von enterischen antimikrobiellen Peptiden (Stockinger et al., 2014a). Die Bildung von Paneth Zellen unterliegt homöostatischen Signalen des angeborenen Immunsystems. So konnten wir zeigen, dass TLR3 und TRIF unter homöostatischen Bedingungen
zur Bildung von Paneth Zellen und damit zur Produktion von antimikrobiellen Peptiden beitragen (Stockinger et al., 2014b).
Nach Sekretion finden sich antimikrobielle Peptide angereichert in der Mukusschicht wieder. Sie bilden dort eine physico-chemische Barriere, die die Penetration von Bakterien hemmt. Darüberhinaus inhibieren sie auch die Stimulation durch mikrobielle Liganden und ermöglichen so die Elimination von Bakterien in der Abwesenheit einer Immunstimulation und tragen zur mukosalen epithelialen Homöostase bei (Meyer Hoffert et al., 2008; Dupont et al., 2014).
In einer systematischen Studie zur Expression verschiedener antimikrobieller Peptide während der postnatalen Entwicklung konnten wir zeigen, dass die bekannten enterischen Peptide, -Defensine und CRS Peptide erst zwei Wochen nach Geburt in signifikanten Mengen produziert werden. Während der ersten zwei Lebenswochen schützt ein Peptid einer anderen Familie, das cathelicidin-related antimicrobial peptide (CRAMP) das neugeborene Darmepithel. Dieser Wechsel des Peptid-Repertoires könnte wichtig sein, um die Etablierung der Darmflora beim neugeborenen Wirt zu ermöglichen (Menard et al., 2008).
Hornef MW, Putsep K, Karlsson J, Refai E, Andersson M. Increased diversity of intestinal antimicrobial peptides by covalent dimer formation. Nat Immunol. 2004 Aug;5(8):836-43.
Ménard S, Förster V, Lotz M, Gütle D, Duerr CU, Gallo RL, Henriques-Normark B, Pütsep K, Andersson M, Glocker EO, Hornef MW. Developmental switch of intestinal antimicrobial peptide expression. J Exp Med. 2008; 205(1): 183-93.
Meyer-Hoffert U, Hornef MW, Henriques-Normark B, Axelsson LG, Midtvedt T, Pütsep K, Andersson M. Secreted enteric antimicrobial activity localises to the mucus surface layer. Gut. 2008; 57(6): 764-71.
Stockinger S, Albers T, Duerr CU, Ménard S, Pütsep K, Andersson M, Hornef MW. Interleukin-13-mediated paneth cell degranulation and antimicrobial peptide release. J Innate Immun. 2014;6(4):530-41.
Dupont A, Kaconis Y, Yang I, Albers T, Woltemate S, Heinbockel L, Andersson M, Suerbaum S, Brandenburg K, Hornef MW. Intestinal mucus affinity and biological activity of an orally administered antibacterial and anti-inflammatory peptide. Gut. 2014 May 7. doi: 10.1136/gutjnl-2014-307150.
Stockinger, S., Dolowschiak, T., Duerr, C.U., Pott, J., Eibach, D., Bäckhed, F., Suerbaum, S., Brugman, M., Hornef, M.W. TRIF determines intestinal epitelial homeostasis but is compensated during mucosal repair. J. Immunol., 2014 (in press)